
The Natural Law of free Evolution – Basic Mechanism and Foundation of the Ecosystem

The ecosystem of earth contains a constant, which is called the “natural law of free evolution” here. It identifies that all lifeforms always adapt, with the evolutionary change of the characteristics, from generation to generation, primarily for their own lasting advantage and benefit. Another species manipulating the adaptation of following generations to its own primary benefit is not sustainable and, therefore, has not emerged stably anywhere in the evolution of ecological structure. And that is why, in the entirety of living nature – apart from our own influences – not a single example has ever been found where any feature of a species has evolved for the exclusive benefit of another. Our civilization, however, has aligned itself with artificial breeding, which is diametrically against this central natural law of free evolution. It has built, thus, almost its entire existence on a foundation which cannot function at all sustainably. Realizing this fact may be extremely hard and painful. But the alternative, that is to continue to bury one’s head in the sand, so to speak, will have – and is actually already having – much worse consequences.
The “natural law of free evolution” is the core mechanism in Charles Darwin’s theory.
The “natural law of free evolution” was central to Charles Darwin’s theory, although he never termed it that way. Several times he wrote that his entire “theory would be destroyed” if somewhere in nature even one single example was discovered where characteristic of any form of life had developed for the exclusive benefit of another species. What follows are some corresponding original quotations from his main work, “On the Origin of Species”:
“Natural selection cannot possibly produce any modification in any one species exclusively for the good of another species; though throughout nature one species incessantly takes advantage of, and profits by, the structure of another. (…)
If it could be proved that any part of the structure of any one species had been formed for the exclusive good of another species, it would annihilate my theory, for such could not have been produced through natural selection. Although many statements may be found in works on natural history to this effect, I cannot find even one which seems to me of any weight. (…)
Natural selection will never produce in a being anything injurious to itself, for natural selection acts solely by and for the good of each. No organ will be formed, as Paley has remarked, for the purpose of causing pain or for doing an injury to its possessor.”
Charles Darwin, On the Origin of Species, 1866 [1] External link : Darwin Online
Darwin was obviously extremely sure of the issue. And since he was never seriously disproved, his quotation is sufficient proof of the existence of the natural law of free evolution. To this day, despite millions of empirical studies of often extremely complex symbiotic and parasitic relationships between viruses, microorganisms, fungi, plants and animals, not a single example has been found of any feature of any species, no matter how tiny, that has evolved for the exclusive benefit of another species. There is a firm reason for this, which becomes clear when considering the structures of genomes and the processes of the development of living beings in the course of evolution.
The absence of intergenerational manipulation in nature can only be due to the fact that it does not work sustainably
One thing that can be immediately excluded is that lifeforms renounce the “breeding” of other species because they obey the “natural law of the free evolution” in the sense of any “morality.” If it worked sustainably, then the kingdoms of the unicellular organisms would have already been worlds of intergenerational manipulation and enslavement for billions of years and there would be no “voluntary” limit as to the degree of these manipulations.
For example, horizontal gene transfer between bacteria is virtually an all-day business in nature and has been for billions of years. And most viruses are highly specialized in manipulating the genome of their hosts in an extremely targeted and precise manner. This targeting and precision often proceeds in much higher dimensions of complexity than our relatively primitive breeding or genetic engineering. However, the viruses only manipulate the genome of the host as an individual – they do not interfere in the germline of the host species.
But why are they not doing it? Why did the evolutionary process, which over billions of years developed such inconceivable complex things like animals with eyes and brain, not create viruses or bacteria that are somehow interfering into the evolution of other species to gain advantages from it?
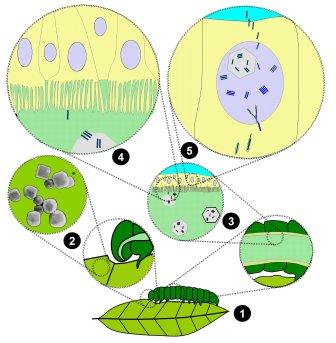
A vivid example for understanding that the lack of “breeding” in nature has nothing to do with lack of theoretical possibility, complexity or purposefulness is the reproductive strategy of viruses of the family Baculoviridae, which has existed for about 300 million years: They manipulate the genetics and neuronal system of caterpillars in an extremely precise way so that they – contrary to their healthy behaviors – crawl up to the highest peaks of the host tree. In addition, the virus disables their feeding limits, causing the caterpillars to become abnormally “fat” and bloat with virus-saturated fluid until they burst, creating a trickling mist of viruses that in turn infects the healthy caterpillars in the lower regions of the plant. This highly complex manipulation strategy, consisting of several extremely complex and targeted steps, is precisely laid out in the genome of the viruses and would not be reproducible by a human breeder or genetic engineer. But never has any virus been discovered that uses these extreme potentials to manipulate the characteristics of the subsequent generations of other species.
The complete absence of any intergenerational manipulation between different species throughout nature allows only one possible conclusion: namely, that it cannot work sustainably. And an reflective analysis of the mechanisms surrounding parasitic relationships reveals the crucial reason: the manipulator would always cause a weakening of the manipulated lifeform if it is not selected primarily for its own benefit. Only when naturally selected by all the many different environmental influences in existence, without targeted manipulation by another lifeform, can a species receive its lasting stability within the constantly changing structure of those environmental influences.
Manipulative intervention in the evolution of a species leads, without exception, to its weakening against the sum of environmental influences
It is true that a manipulator could gain advantages over several generational sequences of an intergenerationally manipulated life form, e.g. by gaining more food. The possible number of successions is variable. But in any case, the manipulated side would “slip between the fingers” of the manipulator from the first of these generations, because it is always weakened in some way by the omission of natural selection for its own advantage in relation to the innumerable interacting and, thus, uncontrollably complex influences of the ecosystem.
As this process proceeds steadily, the advantages of the now specialized manipulator will at some point inevitably turn into disadvantages for its own stable permanence. Since the manipulator had to specialize, it has become dependent on increasingly weakened lifeforms and has, thereby, approached the end of an evolutionary impasse from the beginning. And there, at this end, its own genetic lineage must virtually burn out, like a shooting star.
It is probable that, with the possibility of an exact retrospective observation of earth’s life history, numerous such “shooting stars” could be found. These would be, for example, viruses that had intervened in the evolution of a species of bacteria, in order to make it, in some way, more docile for their descendants. But since the new bacterial generations became rapidly weaker as a result of the manipulation, as far as their permanence in the total structure of the environmental factors is concerned, they died out again, consequently leading to the extinction of the viruses specializing in their use. It is, therefore, the natural selection itself which has ensured that no intergenerational manipulative relationships developed in living nature.
The complexity of each genome is so extremely high that it is uncontrollable from the outside
The actual mechanical basis of the “natural law of free evolution” is easy to identify and can be proved mathematically. It consists in the fact that the information storage capacities of the genome of every lifeform, down to the relatively “simplest” microorganisms and even viruses, are allowing such an extremely high complexity of the information contained that it gradually nears infinity. And these storages existing today have filled up with information over the evolutionary process, which has been going on for several billion years, in order to form organizations capable of surviving, as sustainably as possible, the practically infinitely complex and constantly changing pressure of environmental factors.
At first sight, the mechanical basis might appear to be quite simple: In all lifeforms, the “hereditary information” is stored in a sequence of always identical molecules of only four different nucleotide bases. These are adenine, thymine, guanine and cytosine, which are abbreviated as A, T, G and C (note: RNA viruses excluded here for simplification). The molecules are held in pairs (base pairs) by various auxiliaries and they are arranged in long filaments, which are wrapped into space-saving packages. The whole mixture of nucleotides and auxiliaries is called “deoxyribonucleic acid,” abbreviated to DNA. In every single living cell, there is one or more such package, which are called “chromosomes.” The totality of the genetic material is called the “genome,” and in multicellular organisms this is identical in every single body cell of the individual.
Now, as far as the contained information itself is concerned, it can be simply thought of as codes that lie within the variable sequences of A, T, G, and C. These codes are then used by further cellular organs. They are “read out” as required and then translated into various functions. The best known of these is that of the formation of variable proteins. The physical characteristics of a living being basically consist of many different such proteins, and their respective blueprint is, thus, encoded within the genetic material. In a very simplified and abbreviated way, the total code of the genome could be represented as follows:
AGGATCCTACGTAGTGACCGTTGTCTAAGTCGACTTGAT
In order to recognize that we are really dealing here with a practically unlimited potential of complexity, or one that is nearing infinity, the formula with which the so-called “variation with repetition” is calculated in mathematics is suitable. Written out, it reads simply like this: (n^k). The number of varieties of available nucleotides, i.e., A, T, G and C, becomes “n” (n=4) and the placeholder for the length of the series is “k”. With this, it is now quite easy to look up how the result changes to the possibilities of variation when the length of the genome, k, is increased stepwise. Following are two results:
n = 4
k = 10
(4^10) = 1.048.576
n = 4
k = 200
(4^200)=
258.224.987.808.691.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000
The result at k=10, with a little over one million and, thus, seven digits is not particularly spectacular. But when increasing it 20 times to k=200, suddenly an inconceivably gigantic number with almost 100 digits appears. This number can be written as 258 x 10^93 and might seem rather unspectacular then. There are even names for such large numbers. In this case, it would be 258 quintillion.
The number of all atoms of the universe is miniscule compared with the number of possible variations in even the smallest genomes
What it is really about here, however, can be guessed rudimentarily if one now looks up what is estimated by astrophysicists as an upper limit in another context: namely, the estimated maximum number of all atoms of the universe. This really means the totality of all atomic particles of which the entire universe, with all its galaxies and celestial bodies included. It has to be noted that even the atoms within a single drop of water are far too huge for us to comprehend. Estimates by astrophysicists for the entire universe range between 10^84 and 10^89. So even the highest assumed number would, therefore, fit about 2.5 million times into the calculated number of possible variations of the fictitious genome with 200 base pairs.
But now something much more spectacular comes into play: There is no genome in living nature that is even nearly as “tiny” as just having 200 base pairs. The smallest bacterial genome discovered to date was found in the leaf flea endosymbiont Carsonella ruddii, with almost 160,000 sequences of A, T, G, and C. If one would now fill the formula with k=160,000, then the result would make the estimated total number of atoms of the universe disappear towards zero by comparison.
No mutualism has anything to do with breeding manipulation – real mutualism can only emerge in free evolution
The complexity of the information stored in each genome, nearing infinity, makes successful steering it from the outside absolutely impossible, at least in the sense of a sustainable process. Only if a form of life is selected from generation to generation, according to its own advantage under automatic consideration of all environmental influences, can it remain stable in the long run. This also applies to the emergence of so-called “obligate mutualism.” The term is used for symbiotic relations in which two or more species have mutualistically adapted to each other to such an extent that they could not survive independently without each other. Well-known examples include lichens, which are the result of an obligate mutual symbioses between fungi and bacteria that are so intertwined that, when viewed from the outside, they appear to be a single organism.
Many bacteria living in our human digestive tract are also obligatory mutual partners of our body. We need them, for example, to break down nutrients, regulate acids, or ward off harmful microbes. However, these mutual partners don’t do this because our body has “bred” them into this form of existence. Rather, over millions of years of evolutionary selection, their characteristics have formed primarily for their own benefit. They are helping us in many ways because our body represents their habitat; it is their “home” and – in terms of sustainability – should also be the home of their offspring. That is why their selection to their own advantage primarily is automatically connected with secondary benefits for ourselves as human individuals.
Supposed breeding relationships in nature are always based on distortions or errors
Sometimes, obligate mutualisms superficially look as if there is such a thing as “agricultural” breeding relations after all. However, these are – without exception – always fallacies. They are especially common in the literature surrounding some species of leafcutter ants and umbrella fungus. The lifeforms involved have adapted to each other over probably about 70 million years, to such an extent that both the ants and the fungus could no longer exist independently. Because the symbiosis takes place in such a way that the fungus, which has always lived underground, is supplied with nutrients by the ants and, in turn, produces small nodules that form ideal food for the ants, it is incorrectly interpreted that the ants must have deliberately bred the fungus, just as “civilized man” has done with numerous cereals, fruits and vegetables. But there is no truth at all in this assumption.
The obligatory mutualism between leafcutter ants and the fungus (which also involves some bacteria species as additional mutualistic partners) is one of the most researched ecological relationships ever. There are thousands of studies, and many scientists have invested large parts of their overall work here. But never has a single trait of either the fungus or the ant been shown to have evolved for the exclusive benefit of the other and through intergenerational manipulation of the same. So, the fungus was not manipulated by the ants by “artificial selection” to form the nodules, but it has evolutionarily “noticed” that this effort brings it advantages because it can attract the ants and bind them to itself.
Conversely, however, the ants were not selectively forced by the fungus to align themselves in such a way that they supply it with nutrients, which they do by producing a plant-based paste that precisely fits to the needs of the fungus. Each step of their own adaptations was also always primarily for their own benefits, not for those of the other side. This is why the development of genuine and stable obligatory mutualism needs a very long time, which usually runs over several or even tens of million years. And it is always certain species that adapt to each other.
Not even one of the “useful organisms” in agriculture of man is a mutual symbiotic partner
In order to recognize that an obligatory mutualism in nature can only have superficial similarities with the breeding actions of today’s humans towards “useful animals” and “useful plants,” three clearly identifiable differences can be found:
1. In a true obligate mutualism, only particular participants are specifically aligned with each other. When excluding the microorganisms involved, it is mostly a matter of only two or, at most, three species.
2. The physical and other alignments of the characteristics of the partners relevant for the obligate mutualism mostly run over several millions of years.
3. In a true mutual symbiosis, no characteristics can be found on either side that have been produced by the other side through successful manipulative interference in their evolution for their own primary benefit.
Since these three crucial points do not apply in any single case of the agricultural relationship between man and “useful animals” and “useful plants,” no genuine mutualistic symbiosis partnerships exist. So what we do with our agricultural breeding activities are not mutualistic but parasitic. The “farm animals” and “farm plants” have not gained any primary and certainly no sustainable benefit or advantage through selection for our benefit in terms of their own sustainable permanence.
Their numbers might have increased due to our agricultural activities, but they have – within a tiny period of time by evolutionary criteria – become so weak in the face of countless and extremely complex interacting environmental influences, that most of them would immediately die out as soon as our supports was withdrawn. This extreme weakening process has happened over the course of just millennia and is has nothing to do with mutualism.
The intensive agriculture of the present marks the end of the evolutionary impasse
The weakening and genetic impoverishment of “useful” plants and animals has accelerated sharply, especially in the last phase of intensification of agriculture for the purpose of creating ever more benefits for man. What we can now observe in the fields of crop monocultures or in the halls of the livestock industry are organisms that are so weak in the face of the many components of environmental pressure that they have to be supported at great expense. Therefore, they need increasing support such as water, fertilizers, insecticides, herbicides, fungicides, antibiotics, antivirals and many other aids.
The phase of global intensification in agriculture, starting in the middle of the last century, has led in parallel to a short but sharp increase in the amount of food generated and, subsequently, to an explosive increase in the human population, from about three billion in 1950 to close to eight billion in 2020. In the next few years of this stalemate, the inevitable further weakening of agricultural organisms will lead to an escalation in the use of chemical support to sustain them. This will subsequently put further pressure on the ecosystem and, not least, allow parasites to select themselves into new extreme forms. Altogether, it can be drawn as a logical conclusion that a collapse of this completely unnatural and unstable structure lies close ahead.
The “natural law of free evolution” is rarely considered in today’s natural sciences, despite its central importance, as shown in earlier statements by Charles Darwin, for the same reason as freedom as the central regularity of nature is repressed. But the natural law of free evolution is the mechanical basis for freedom; as there is no lasting possibility of mutual breeding, every living being is born free, in the sense of the word, and enslavement relationships between different species have remained impossible since the beginning of life.
Conclusion
The “natural law of free evolution” has always ordered life on earth and it is based on the externally uncontrollable complexity of all genomes. It formed the basis of over 500 million years of history of higher life on the planet with its enormous variety. Only in free evolution such can arise and remain stable. Moreover, this law of nature is the foundation of the central regularity of freedom, because it excludes the possibility of lasting enslavement between different species. The very broad alignment of man against these central laws of nature and regularities from the “Neolithic Revolution” onwards has led civilization into an evolutionary impasse. Numerous parallel culminations of the present day suggest that its end is either imminent or that it has already been reached.
Sources:
[1] Charles Darwin, On the Origin of Species: fourth British edition (1866), page 241
http://darwin-online.org.uk/Variorum/1866/1866-241-c-1859.html